


Ancestral Health vs. Antagonistic Pleiotropy?
Why Nick Hiebert is wrong in using antagonistic pleiotropy to refute the ancestral health framework.

I recently had a debate on Twitter Spaces with Nick Hiebert on the value of the ancestral health framework. I will publish this soon with my reflections.
Here, however, I want to make an independent analysis that focuses narrowly on one of Hiebert’s claims, derived from materials of his and others I consumed after we finished the debate.
Specifically, the claim that the principle of antagonistic pleiotropy suggests artificially manipulated foods are more likely to promote longevity than ancestral foods.
This claim is made in Hiebert’s article, Should Modern Humans Eat Like Hunter-Gatherers?
Hiebert revised this article today in response to a private discussion we had. Google has a cache of the original argument as last updated in January, 2021 here.
I quote from the original first:
So, the question ends up being: how long after reproductive age does natural selection still robustly apply to human beings? Surely selection pressure doesn't end at reproductive age, because human children need human adults to raise them and care for them. But does selective pressure exist to a meaningful degree for those in the age ranges that typically associate with life-threatening chronic disease? Some scholars have attempted to estimate the force of natural selection as a function of age [6]. . . . our best estimations suggest that the forces of natural selection rapidly wane down to nil shortly after sexual maturity, which would be approximately 16 to 17 years of age. Around the ages of 30 to 40 is when humans likely enter the "selection shadow", which is the zone wherein natural selection no longer robustly applies. . . .
It is also imperative to mention that foods to which we are adapted might actually be more likely to be harmful for us as we age. This is because of a concept in evolutionary biology called antagonistic pleiotropy, which is the most widely accepted explanation for the evolutionary origin of aging [12][13]. The theory of antagonistic pleiotropy essentially posits some genetic adaptations can trade long-term health for short-term reproductive success. However, it also turns out that most genetic adaptations are antagonistically pleiotropic [14]. Since adaptations to foods are no more or less genetic than any other adaptations, we can infer that most adaptations to food are also likely to be antagonistic pleiotropic. . . .
Without scientific evidence to help inform our attitudes toward the relative health value of novel foods, the health value of those foods remains is a black box. Due to the principle of indifference, we have no particular reason to suspect that truly novel foods will be either beneficial or detrimental. However, if it can be demonstrated that both an ancestral food and a novel food have the same thriving potential during the reproductive years, we can actually infer that the novel food is to be favoured.
Novel foods do not belong to the domain of foods that have the potential to be antagonistic pleiotropic, since we did not evolve consuming them. Conversely, the ancestral food in our scenario is likely to be antagonistically pleiotropic.
Hiebert makes four serious errors here:
He underestimates the force of selection in later ages when applied across extremely long amounts of evolutionary time.
His argument that antagonistic pleiotropy implies a health advantage of novel foods over ancestral foods gets the significance of adaptation completely backwards. This is because he treats the lack of adaptation to a food as good for longevity when in fact it is bad for longevity. In fact, his argument has been mathematically and experimentally falsified by evolutionary biologist Michael Rose. Or, rather, Rose showed the precise opposite many years before Hiebert ever got it backwards.
His quantitative claim traces to a reference that is irrelevant. It is a paper showing that pleiotropic genes are more likely to be conserved than non-pleiotropic genes in yeast. He seems to be confusing a quantitative claim about pleiotropy itself (a gene having more than one function) with a quantitative claim about antagonistic pleiotropy (a tradeoff between fertility and longevity).
He conflates genetic adaptations to foods with the foods themselves. Even if it were true that the majority of food-related adaptations were antagonistically pleiotropic, this wouldn’t tell us anything about the amount of foods that have this property.
Is Natural Selection Significant for Late Ages?
Natural selection is driven by those who remain healthy before and during their window of fertility being more likely to leave offspring than those who fall ill or die before or during that same window.
However, this does not mean that natural selection has no effect on health in late life.
First, as evolutionary biologist Michael Rose reviewed here, the end of the window of fertility is defined as the latest age of reproduction in a population’s evolutionary history. Even now, we have women getting pregnant as late as age 74 and men getting women pregnant as late as age 94. Granted, it is possible none of our ancient ancestors had children this late, but we know it is physically possible so we have to regard it as the plausible end of the evolutionary window of fertility.
Since many genes do more or less the same thing in men and women, some of the genes that are selected for by men siring children in their 90s will also benefit women in their 90s. Thus, the absolute end of the evolutionary window of fertility for humans really extends into the 90s.
As also reviewed by Rose, because some genes that promote health during the window of fertility also promote health after the window of fertility, the health-promoting force of natural selection reaches an end somewhere after the last age of reproduction in a population’s history. Thus, in humans, there will be at least some effect of natural selection right through the hundredth decade of life and perhaps even later.
Because the window of fertility is mostly concentrated at earlier ages than the maximum, and because most pregnancies have occurred toward the earlier part rather than the later part of that window, quantitatively natural selection is promoting health primarily at the earlier part, is doing little at the later part, and is having exceedingly small effects after the window is over.
Adaptations that benefit health before the end of the fertility window accumulate much more rapidly than those that benefit health after that window. However, adaptations that promote health after that window still accumulate. They just accumulate very slowly because the force of natural selection is so weak.
Hiebert shows this graph from this paper:

He uses a somewhat different version that I do not see anywhere in the reference:
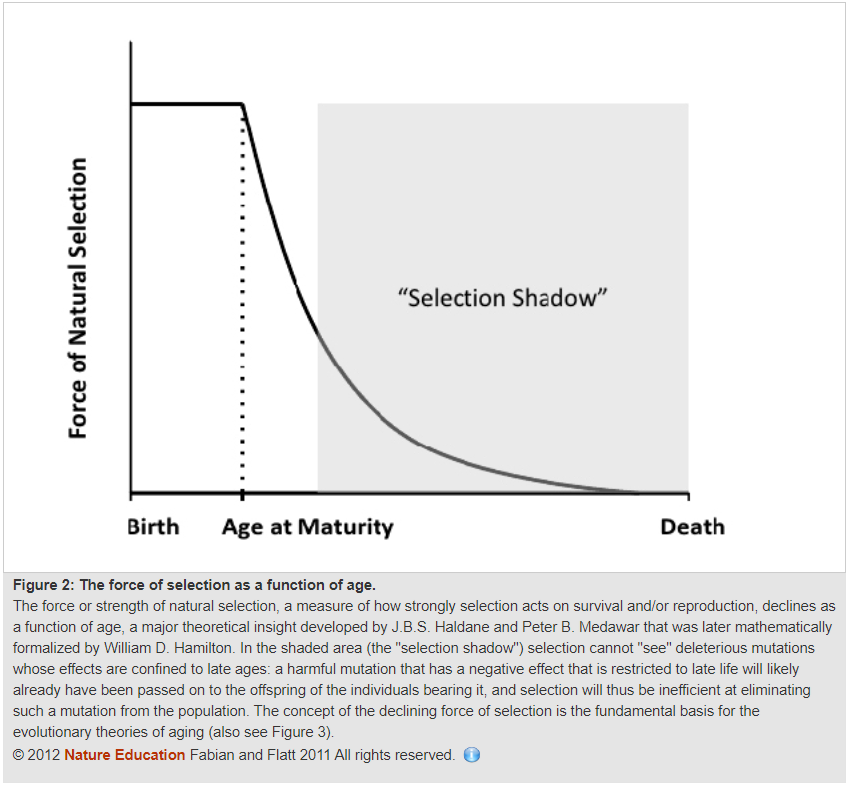
In reference to this graph, he writes the following:
Applying the above graph to human beings, our best estimations suggest that the forces of natural selection rapidly wane down to nil shortly after sexual maturity, which would be approximately 16 to 17 years of age. Around the ages of 30 to 40 is when humans likely enter the "selection shadow", which is the zone wherein natural selection no longer robustly applies.
In the top graph, the one I find in the paper he referenced, the solid line represents the force promoting survival, and the dashed line represents the force promoting the ability to reproduce.
“Nil” means “zero,” yet the top graph shows the survival-promoting force of evolution declining only about 60% by the time the “selection shadow” is entered, and it remains non-zero until nearly the end. The second graph is similar.
Hiebert does not define “robust” but he seems to imply that natural selection cannot be relied on to do much of anything in late life.
These are both gross underestimations of what natural selection can do after the main window of fertility. Its effects do not reach anything remotely close to “nil” shortly after sexual maturity, and they don’t even reach nil at the average age of death.
The average age of death is earlier than the latest possible age of reproduction in humans, so even if the force of natural selection is exceedingly weak, it still has the ability to produce meaningful longevity benefits over exceedingly long periods of evolutionary time.
Thus Hiebert’s first error is confirmed: He underestimates the force of selection in later ages when applied across extremely long amounts of evolutionary time.
This brings us to Michael Rose’s experiments.
Antagonistic Pleiotropy Means Novel Foods Hurt Longevity More Than Ancestral Foods
Antagonistic pleiotropy refers to some genes having a beneficial effect early in life and a harmful effect late in life. In other words, they trade longevity away to gain fertility.
Natural selection has the ability to allow or even select for such genes, because it acts so powerfully early in life and so weakly late in life.
Hiebert claims that this principle renders novel foods more likely to promote longevity than ancestral foods:
The theory of antagonistic pleiotropy essentially posits some genetic adaptations can trade long-term health for short-term reproductive success. However, it also turns out that most genetic adaptations are antagonistically pleiotropic [14]. Since adaptations to foods are no more or less genetic than any other adaptations, we can infer that most adaptations to food are also likely to be antagonistic pleiotropic. . . Novel foods do not belong to the domain of foods that have the potential to be antagonistic pleiotropic, since we did not evolve consuming them. Conversely, the ancestral food in our scenario is likely to be antagonistically pleiotropic.
This is Hiebert’s second error: he gets the significance of adaptation completely backwards because he treats the lack of adaptation to a food as good for longevity when in fact it is bad for longevity.
In 2018, evolutionary biologist Michael Rose gave a presentation on this topic at the Ancestral Health Symposium (thanks to Jevan on Twitter for making me aware of it):
Rose doesn’t use the term “antagonistic pleiotropy” in his talk, but he showed experimentally in 2002 that the differences in longevity among the fruit flies he used in his experiments were likely driven by antagonistic pleiotropy.
Because fruit flies have such short lifespans, Rose was able to adapt them to diets over more than a thousand generations, the equivalent of 20-30,000 years of human evolution.
He was therefore able to study experimentally how genetic adaptations to foods affect the longevity response to ancient, recent, and novel diets.
His results line up well with his mathematical predictions and demonstrate the complete opposite of Hiebert’s argument.
Here are the diets:

When Rose’s team first obtained the flies, they had already been feeding on rotten apples for centuries. This represents their ancestral diet, much like a human “Paleo” diet. Then they switched them to bananas, and kept them on bananas for over 1,000 generations. This represents 20-30,000 years of human evolution, considerably more time than we have spent adapting to agricultural diets.
Once they had an “ancestral” diet representing the vast majority of the flies’ evolution and an “evolutionarily recent diet” representing the human equivalent of 20-30,000 years of human adaptation, they could compare the longevity response of each to an “entirely novel diet” made of oranges.
Mathematically, their success as a population is measured as their probability of survival (px) times the numbers of eggs laid by each female (mx).
Since natural selection acts rapidly on health up through the window of fertility and very slowly thereafter, their model assumes that the flies had enough evolutionary time to acquire longevity-promoting adaptations to the ancestral diet, but only enough time to acquire fertility-promoting adaptations to the evolutionarily recent diet, and no time to acquire any useful adaptations to the novel diet.
Thus, their model predicts that the ancestral diet promotes health across the lifespan, while the evolutionarily recent diet promotes health primarily during the fertile years, and the novel diet does not even have the fertility benefit.
This is shown below. In the predictive model, the entirely novel diet (dotted orange line) underperforms the other two diets through the window of fertility. After the window of fertility, the evolutionarily recent diet (blue line) fails to support longevity any better than the novel diet, and only the ancient diet (dotted red line) outperforms.

Experimentally, the data are noisier than the model, but Rose and his team showed that the evolutionarily recent diet outperformed the novel diet at early ages:

And the ancient diet outperformed the evolutionarily recent diet in older ages:

Thus, the experimental data are noisy but the trends observed support the predictive model.
If these data are driven in part by antagonistic pleiotropy, it would appear that shorter periods of evolutionary time are more likely to cause the accumulation of antagonistically pleiotropic adaptations, while much longer periods of evolutionary time ultimately favor longevity-promoting adaptations that displace the antagonistically pleiotropic ones or overwhelm them in importance.
The reason that these experiments show the opposite of Hiebert’s argument is that he mistakes being adapted to a food as being bad for longevity, when in fact it is good for longevity.
Hiebert’s mistake is partly located in his first error, in which he dismisses any useful effect of evolutionary forces in late age. He thus underestimates how much beneficial adaptation could occur over very long periods of time, such as those we spent eating ancestral diets.
His mistake, however, is also rooted in his third error: his assumption that most adaptations are antagonistically pleiotropic.
Let’s move on to that error now.
Are Most Genetic Adaptations Antagonistically Pleiotropic? Part 1
First we tackle this error as Hiebert originally had it, prior to today, when he revised it in response to a conversation we had:
The theory of antagonistic pleiotropy essentially posits some genetic adaptations can trade long-term health for short-term reproductive success. However, it also turns out that most genetic adaptations are antagonistically pleiotropic [14]. Since adaptations to foods are no more or less genetic than any other adaptations, we can infer that most adaptations to food are also likely to be antagonistic pleiotropic.
His reference for “it also turns out that most genetic adaptations are antagonistically pleiotropic” is a paper on yeast showing that “the evolutionary conservation of genes and gene sequences positively correlates with the level of gene pleiotropy.”
Pleiotropy simply means having more than one function. All this paper showed is that genes that have more than one function are more likely to be evolutionarily conserved in yeast.
Nick seems to be confusing pleiotropy per se with antagonistic pleiotropy.
The first is simply a gene having more than one function. The second is the principle that natural selection favors trading off longevity to gain fertility. The first does not in any way automatically imply the second.
And it’s in yeast, not humans.
Are Most Genetic Adaptations Antagonistically Pleiotropic? Part 2
Today, in response to a conversation we had, Hiebert deleted the yeast paper and replaced it with an argument about senescence, the deterioration that occurs with age:
[Premise 1]) If gene degradation is an antagonistically pleiotropic trait and most human genes degrade, then most human genes are antagonistically pleiotropic. . . .
[Premise 2]) Gene degradation is an antagonistically pleiotropic trait. . . .
[Premise 3]) Most human genes degrade. . . .
[Conclusion]) Therefore, most human genes are antagonistically pleiotropic. . . .
Essentially, human DNA tends to degrade over time when it doesn't rightfully need to, as evidenced by the existence of biologically immortal organisms. Human DNA repair is also regulated and gene-specific. Given these facts, DNA degradation in humans itself is likely to be adaptive. This assigns every gene in our DNA that degrades over time a single antagonistically pleiotropic trait. Since most DNA in the human genome degrades over time, we can infer that over 50% of genes are antagonistically pleiotropic.
Hiebert’s first premise is wrong, because a gene degrading is not a function of that gene. If broad swaths of the human genome are all victims of an age-dependent decline in DNA repair, this isn’t a pleiotropic function of each of those genes. It is just a process that happens to them.
The principle of antagonistic pleiotropy is that a specific gene has at least two specific functions, at least one of which is good for fertility and at least one of which is harmful for longevity.
Let’s posit a hypothetical gene with a single function, to do X. It doesn’t matter what X is. The gene is monotropic, not pleiotropic. It has one function, not two, or ten. If the gene is expressed, and is vulnerable to DNA damage, and DNA repair undergoes global age-dependent decline, then it will accumulate mutations with age. Is this a “second function” of the gene? No. It’s just losing its singular function.
Hiebert’s second premise is supported merely by his later statement that “Human DNA repair is also regulated and gene-specific. Given these facts, DNA degradation in humans is likely to be adaptive.”
He is trying to argue that DNA degrading in youth has some beneficial effect despite it hurting longevity, thus making it antagonistically pleiotropic.
This is ridiculous. The fact that DNA repair is regulated and gene-specific is far more indicative that repairing genes is adaptive, not that letting them degrade is.
Of course, there is an obvious benefit to sometimes letting genes degrade, and that is that it would conserve energy that is needed for something regarded as more important. I’m open to the fact that there is some antagonistic pleiotropy to be found in DNA repair genes, but Hiebert hasn’t even come close to supporting that simply by pointing out that DNA repair is regulated.
Regardless, his first premise is so faulty that it doesn’t even matter. If DNA repair genes demonstrate antagonistic pleiotropy, it in no way follows that every gene that gets repaired or fails to get repaired somehow mystically becomes a participant in that antagonistic pleiotropy.
Are Most Genetic Adaptations Antagonistically Pleiotropic? Part 3
Having refuted the two different versions of Hiebert’s premise that most adaptations are antagonistically pleiotropic, let us now look at what he concludes from this.
He argues that because adaptations to food are “no more or less genetic than any other adaptations,” that human adaptations to food must be more often antagonistically pleiotropic than not. Antagonistic pleiotropy does not exist because something is genetic. It happens because a specific gene has at least two specific functions, at least one of which helps fertility and at least one of which hurts longevity.
Even if either of Hiebert’s arguments — from the yeast paper, or from the DNA repair syllogism — had shown that most genetic adaptations in total demonstrate antagonistic pleiotropy, it wouldn’t follow that human genetic adaptations to food would show the same proportion of antagonistic pleiotropy as the total.
For example, it wouldn’t follow that human genetic adaptations to food would show the same proportion of antagonistic pleiotropy as all genetic adaptations within the yeast genome simply because human food-related genetic adaptations are just as genetic as yeast genes.
Are Most Genetic Adaptations Antagonistically Pleiotropic? Part 4
Let’s close this out by considering the results of Michael Rose’s experiments discussed above.
If the net effect of exceedingly long periods of evolutionary time was to accumulate adaptations to the ancestral diet that give it longevity-promoting effects, it cannot possibly be the case that the net effect of natural selection is to promote accumulation of food-related genetic adaptations that exhibit antagonistic pleiotropy with a net anti-longevity effect.
Granted, we can’t know the number or proportion of genes promoted that exhibit this trait because Rose’s team never mapped out all the genes involved.
We can say, however, that either the majority of genetic adaptations to the diet were longevity-promoting, or the longevity-promoting adaptations that accumulated were far more important than the longevity-hurting adaptations that accumulated.
It ultimately doesn’t matter if the majority or even most adaptations were for genes exhibiting antagonistic pleiotropy because the net result was a longevity benefit of the ancestral diet. Whether that is because of a greater number of longevity-promoting adaptations or a greater importance of a smaller number of longevity-promoting adaptations is not important.
Thus Hiebert’s third error, his assumption that most adaptations are antagonistically pleiotropic, is definitely not supported by what he wrote in either the first or second version of his article, possibly refuted by Rose’s experiments, and probably false, but ultimately irrelevant.
However, Hiebert gives this quantitative claim a bizarre false significance when he makes his final error: conflating adaptations to foods with the foods themselves.
Conflating Genetic Adaptations With Foods
After falsely assuming most genetic adaptations are antagonistically pleiotropic, then falsely generalizing this false quantitative claim from the total pool of adaptations to the subset of human food-specific adaptations, Hiebert then makes the wildest conflation yet in extending this quantitative claim to the foods themselves:
Since adaptations to foods are no more or less genetic than any other adaptations, we can infer that most adaptations to food are also likely to be antagonistic pleiotropic. . . . Novel foods do not belong to the domain of foods that have the potential to be antagonistic pleiotropic, since we did not evolve consuming them. Conversely, the ancestral food in our scenario is likely to be antagonistically pleiotropic.
Here Hiebert argues that because most evolutionary adaptations to food are antagonistically pleiotropic, most ancestral foods also have this property.
Let us just assume for the moment that the premise is true, that most adaptations to foods are antagonistically pleiotropic. It isn’t, but let’s indulge it.
Does this quantitative claim somehow extend to the foods themselves?
Let’s take an example that Hiebert gave during the debate we had. He offered the example of red meat raising the concentration of ApoB-containing lipoproteins, such as LDL. He said this is a benefit in youth because it prevents fatty liver, but comes at the expense of longevity because it promotes heart disease. Thus, dietary adaptations to red meat and red meat itself are antagonistically pleiotropic.
Without wanting to get into the weeds on how we might see the relation between ApoB-containing lipoproteins and cardiovascular disease differently, and without wanting to get into a detailed disagreement on the relation between red meat and fatty liver, I countered as follows.
Adaptations to raise ApoB-containing lipoproteins were not adaptations to red meat, but rather adaptations to infectious disease. I have written about this here. Since these lipoproteins offer some protection against infectious disease, genes that raise them to levels higher than what would optimize for cardiovascular health were favored prior to the 20th century when infectious diseases were the major causes of death.
That red meat raises them is incidental to this.
If it could be shown that we have genes that prompt us to favor eating red meat, or that we have genes that enhance our ApoB response to constituents of red meat, and that these were selected for during periods of infectious disease abundance in order to elicit the protection of ApoB-containing lipoproteins, then it could be argued that those genes demonstrate antagonistic pleiotropy and were selected because of it.
However, under no circumstance could you say the food itself has been selected for in a way that elicits this tradeoff because human evolution is not what has driven the existence of red meat in the food supply. The existence of red meat animals is driven by their own evolution and their overall place in the ecology.
Thus, even if you could show that genetically ingrained human food-seeking behaviors more often than not trade longevity for fertility, it is a complete non-sequitur to say that foods that existed ancestrally are more often than not making this tradeoff themselves.
And this is, I suppose, why Nick has never attempted to show that foods available ancestrally are more likely to raise ApoB-containing lipoproteins than to leave them flat or lower them.
This is critical for Nick’s argument that artificially manipulated foods are more likely to benefit longevity than ancestral foods because he is starting from a quantitative assumption about the majority of genetic adaptations and making a logical leap to the majority of ancestral foods. His quantitative assumption is likely wrong and ultimately irrelevant, as discussed above, but now consider the problem with his logical leap:
If 75% of genetically ingrained food-seeking behaviors related to ApoB-affecting foods were to lead us to consume a diet that, in net, raises ApoB to higher levels than would optimize for cardiovascular disease, it would not in any way whatsoever follow that 75% of all ancestral foods that affect ApoB raise it instead of lowering it.
In fact, the first point could be true even if this were driven entirely by a voracious appetite for a singular ApoB-raising food, leaving well over 99% of ancestral foods in the ApoB-lowering category.Similarly, and for the same reasons, if 75% of all genetic adaptations to food were bad for longevity, it would not in any way whatsoever follow that 75% of all ancestral foods are bad for longevity.
Again, the first point could be true even if it were driven entirely by a voracious appetite for a singular longevity-destroying food, leaving well over 99% of ancestral foods in the longevity-boosting category.
That is, the proportions of adaptations to foods that demonstrate some property has nothing to do with the proportions of foods that demonstrate that property.
The Final Syllogisms: Finding Safety in Artificial Foods
What does Hiebert conclude on the foundation of the four major errors I identified above?
That artificial foods are more likely to promote longevity than ancestral foods!
[Premise 1]) If a given novel food has non-inferior health value to a given ancestral food in the reproductive window, then such a novel food is likely to be superior to such an ancestral food in the post-reproductive window. . . .
[Premise 2]) A given novel food has non-inferior health value to a given ancestral food in the reproductive window. . . .
[Conclusion] Therefore, such a novel food is likely to be superior to such an ancestral food in the post-reproductive window
He then goes on to define novel foods as including “artificially manipulated foods” and to argue that “artificially manipulated foods have more potential to reduce disease risk than natural foods.”
The first premise in the above syllogism is based on the quantitative claim that ancestral foods are more likely to hurt longevity than novel foods.
This is based on a pile of errors:
Even if it were true that the majority of genetic adaptations to ancestral foods hurt longevity, it wouldn’t follow that the majority of ancestral foods hurt longevity. Hence there would be no basis for the relative likelihood of food from one or the other category hurting longevity.
Even if it were true that the majority of total genetic adaptations in yeast or across all species hurt longevity, it wouldn’t follow that the majority human food-related adaptations to ancestral diets do the same.
It probably isn’t true that the majority of total genetic adaptations are antagonistically pleiotropic, but it’s ultimately irrelevant because the net effect of exceedingly long periods of evolutionary time is to promote adaptations that in net promote longevity on an ancestral diet.
The first premise is also based on the error that not being adapted to a food is good for longevity, when Rose’s experiments showed it is actually bad for longevity.
Conclusion
Ultimately this is all driven by the first error: a lack of appreciation of how much benefit natural selection can bestow upon late-life longevity given exceedingly long periods of evolutionary time.
Obviously, in the end, death always wins. But this is a natural result of the second law of thermodynamics. What deserves explanation is not why we die, but why we even live in the first place, let alone why we live for so long.
A biologically immortal jelly fish doesn’t contradict this. It just has evolved its own way of defying the second law through clever uses of energy.
Certainly selection allows or even promotes some antagonistically pleiotropic genes to accumulate, and these genes may explain a lot about aging.
But the slow, steady force of natural selection over exceedingly long periods of time promotes human longevity in defiance of the inexorable force of the second law driving everything toward meaningless mixtures, and it promotes the accrual of longevity-promoting responses to our ancestral diets.
Disclaimer
I am not a medical doctor and this is not medical advice. My goal is to empower you with information. Please make all health decisions yourself, consulting sources you trust, including a caring health care professional.
Please Show This Post Some Love
Let me know what you think in the comments! And please like the post if you found it valuable, and share the post with others if you believe they too would find it valuable.
Join the Next Live Q&A
Have a question for me? Ask it at the next Q&A! Learn more here.
Subscribe
Subscribe or upgrade your subscription here:
Join the Masterpass
Masterpass members get access to premium content (preview the premium posts here), all my ebook guides for free (see the collection of ebook guides here), monthly live Q&A sessions (see when the next session is here), all my courses for free (see the collection here), and exclusive access to massive discounts (see the specific discounts available by clicking here). Upgrade your subscription to include Masterpass membership with this button:
Learn more about the Masterpass here.
Take a Look at the Store
At no extra cost to you, please consider buying products from one of my popular affiliates using these links: Paleovalley, Magic Spoon breakfast cereal, LMNT, Seeking Health, Ancestral Supplements. Find more affiliates here.
For $2.99, you can purchase The Vitamins and Minerals 101 Cliff Notes, a bullet point summary of all the most important things I’ve learned in over 15 years of studying nutrition science.
For $10, you can purchase The Food and Supplement Guide for the Coronavirus, my protocol for prevention and for what to do if you get sick.
For $29.99, you can purchase a copy of my ebook, Testing Nutritional Status: The Ultimate Cheat Sheet, my complete system for managing your nutritional status using dietary analysis, a survey of just under 200 signs and symptoms, and a comprehensive guide to proper interpretation of labwork.
Oh why oh why did you talk to this nuttyvore? I followed Tucker's analysis of his misunderstandings, and checked his position on TPN. Which was bs and 180 degrees wrong. Falsus in omnibus was my verdict.
My fears realized when I checked his recent tweets (your link). His writing is below bar, by any measure. Dumb and laborious do the most damage, whatever they take up to.
JR
What about the increasing rates of infertility among the youth eating novel foods?
Processed foods are inferior, even in the face of human reproductive resilience.
Look no farther than how food quality affects the quality of breast milk to prove that ancestral foods are in fact superior.