


Sauna vs. Homocysteine
A Self-Experiment for Methylation Health (and Battling Gray Hair)

Many rare disease genes and even some very common genetic polymorphisms hurt enzyme function because the make the enzymes “thermolabile," which means that body heat is “too hot” for the enzyme and denatures it the way more intense heat would denature any other protein.
Often this causes small changes to the shape of the protein that alters its affinity for a nutritional cofactor. The enzyme then becomes more likely to lose the cofactor, and the loss of the cofactor further destabilizes it and makes it permanently denature.
In such cases, maintaining high levels of the nutritional cofactor can increase the stability of the enzyme.
This is true with the infamous MTHFR C677T polymorphism, of which 55% of the world’s population has at least one copy.
Much of the basic research around this enzyme is done in an E coli model, because it is easier to systematically study. This model originally showed that C677T causes MTHFR to thermally destabilize and lose the riboflavin cofactor more effectively, and that both riboflavin and folate can help prevent it from destabilizing.
1.6 extra milligrams of riboflavin, bringing total intake up to 3.2 milligrams per day, abolishes the effect of C667T on homocysteine in humans.
Well, guess what else fixes this polymorphism? According to the E. coli model, heat shock proteins! This suggests that interventions to increase heat shock proteins, such as sauna, could benefit MTHFR.
But hold up — we will see soon that sauna has an acute effect on heat shock proteins, not a long term one.
And what about COLD? If the heat of body temperature is too much for this enzyme, don’t we want to stay cool rather than hot? Wouldn’t sauna hurt, at least temporarily? Should we be cold plunging instead? We will see below that we can actually leverage sauna to cool the body if we do it right.
COMT is another example. This enzyme uses methylation to support mental flexibility and to prevent excessive rumination. About 25% of the population is homozygous for the “slow” version (Val158Met) and 50% is heterozygous. Slow COMT is associated with anxiety and depression, but with good academic performance. The “slow” version is thermolabile, and is stabilized by S-adenosyl-methionine (SAM or SAMe), the universal methyl donor.
The rs1126809 R402Q allele of tyrosinase (TYR) is yet another example. This is likely a major driver of gray hair. It is very common in people with European ancestry, where just under 40% of people are heterozygous and 7.6% are homozygous. It is not found in many Asians. It is found in American Blacks and Latin Americans, but not in African pygmies or Australian Aborigines, leading to speculation that it arose in Caucasians and spread to other groups through admixture. Thermolability causes body heat to loosen its hold on the copper cofactor. Being copper deficient probably makes this worse, but, due to the toxicity of copper overload, it is not clear whether it is safe to supplement with extra copper the way you want to get extra riboflavin for MTHFR.
One of the most fascinating cases of thermolability is the cystic fibrosis mutation, a deletion in CFTR at F508 (rs113993960), which only 0.6% of the world’s population carries. This mutation makes the protein responsible for preventing cystic fibrosis thermolabile. The deletion at F508 is the most common cystic fibrosis mutation. Generally cystic fibrosis cohorts are 50% homozygous for this deletion, but another 40% are heterozygous and may have a less common mutation alongside it.
The mean annual temperature where a cystic fibrosis patient resides is a predictor of lung function, such that each 10F (5.6C) increase in temperature is associated with a 3% loss in lung function. When this mutation is engineered into mice, taking their cells and incubating them at 26C (79F), instead of body temperature, leads to between 50% and 100% restoration of function. In another study, mice with this deletion had their temperature set point chemically lowered with a chemical injection, then were kept at air temperatures of 15-17C (59-63F). Whole-body cooling partially reversed much of the pathology, improving mucous cell presence and salivary flow, and moderately reducing mortality. Altogether this does not support using whole-body cooling as a cure for cystic fibrosis but it does support environmental temperature as a causal modifier of the disease severity.
Whether we can “cure” the common polymorphisms in MTHFR, COMT, and TYR is a different story. These are, first of all, common traits in the population and not diseases requiring a cure. But the fact that a little extra riboflavin abolishes the impact of C677T on methylation as judged by homocysteine levels suggests that totally abolishing the effect of any of these polymorphisms could be in reach.
MTHFR C677T and COMT Val158Met correlate positively with latitude. This and the concentration TYR R402Q in Europeans suggests that the cold climates humans experienced as we migrated out of Africa and into the ice age-dominated global north could have allowed the thermolabile variants to spread. This, then, suggests that for those of us with these common polymorphisms, our mental health, our blood cell synthesis, the development of healthy babies, and our graying hair are all screaming at us to get into the cool temperatures that our bodies were designed for.
In trying to hack our way out of the fates imposed by these variants, we need to be careful to distinguish between environmental temperature, acute temperature stress from saunas and cold plunges, and the counter-productive drop in body temperature that can be caused by a lower metabolic rate.
We do not want to do anything that would lower ATP levels, because ATP fuels the activity of heat shock proteins, is needed to make methylfolate, and is needed to activate methionine to S-adenosyl-methionine, the universal methyl donor. So we don’t want to just tank our metabolic rate to improve the function of these enzymes.
The protocol I advocate for this could be called sauna hard and stay cool.
This is educational in nature and not medical or dietetic advice. Any products linked herein may be affiliate links. See terms for additional and more complete disclaimers.
Sauna Hard and Stay Cool
The sauna hard and stay cool protocol is as follows:
Sauna 30-60 minutes anywhere from once every two days to twice a day, at 80-100C (176-212F) and 30-40% humidity.
Men should keep their testicles wrapped in ice packs in the sauna.
Keep your house as cool as you can tolerate, down to 15.5 C (60 F). Stay at this temperature as much as possible throughout the day, every day. Do not bundle up to stay warm — get used to the discomfort and learn to find it comfortable.
Cold plunging is totally optional and not an important part of the protocol.
GET MEDICAL CLEARANCE: Make sure you are in healthy enough shape to handle intense sauna usage. You could suffer heat stroke or if you have a severe thermolabile genetic mutation sauna could induce metabolic harm. Work your way up to the heat and humidity recommended here to make sure you tolerate it. Make sure to stay hydrated and get your salt and other electrolytes replaced.
A Self-Experiment For You
Use the sauna hard and stay cool protocol to test blood levels of homocysteine, the methylation index on the Dutch Complete test (which measures the methylation of estrogen by COMT), or the grayness of your hair.
It should take at least seven sauna sessions to reach maximal adaptation, and the adaptations can likely be lost within 2-3 days.
A reduction in homocysteine or an increased Dutch methylation index should be apparent within a week of this protocol.
Hair graying is impacted toward the middle of a very long cycle of hair growth. Treatments that induce re-pigmentation take at least one month, often six to twelve months, and sometimes up to two years.
For homocysteine and the methylation index, the timescale on which you would expect this to work could be short enough that you could perform a rigorous randomized controlled trial on yourself. Briefly, do a series of measurements to verify that the effect appears to be maximized at one to two weeks (or whatever you find). Wherever the effect is maximal, this is the duration of each experiment. Use a random number generator or some other method to randomize periods of that length to either sauna hard and stay cool or control. Do at least three measurements of each. Further guidance can be found in How to Do a Proper Self-Experiment, and Why Your “N” Doesn’t Technically Equal “1”
You don’t have to do a randomized experiment. You can just try it and see if it seems to work.
For hair graying, you need to persist for at least a month and possibly much longer, so in this case you want to just stick with it if you like it and feel good on it and wait to see if it works.
Keep your diet and exercise regimen as consistent as possible across the length of your experiment.
Post Your Results Here!
Once you have results, come back here and post them in the comments!
The Justification for the Protocol
Now we turn to the justification for the sauna hard and stay cool protocol.
In this section we will learn the following:
Your environment determines your body temperature.
Long-term adaptation with extreme cold exposure can lower body temperature further, but requires high volumes of discomfort that go beyond any cold plunge protocol that has been studied.
Sauna is the most effective way to lower resting core body temperature while benefiting rather than hurting the metabolic rate.
Heat-shock proteins seem to be a temporary adjustment that become irrelevant once the long-term adaptations to sauna settle in.
The “stay cool” part of this should increase your metabolic rate, an extra boost.
In This Article:
Your Environment Determines Your Body Temperature
Extreme Cold Exposure Causes Long-Term Decreases in Body Temperature That Have Never Been Replicated in Cold Plunge Studies
Sauna Lowers Resting Body Temperature
Exercise Can But Doesn’t Always Replicate Sauna Heat, And Never Replicates the Decrease in Resting Temperature
Why Do Exercise and Sauna Differ?
What About Heat Shock Proteins?
Will Sauna Wreck Your Energy Metabolism?
Caveat: Sauna Can Probably Wreck Your Sperm Without Ice
Your Environment Determines Your Body Temperature
While we are warm-blooded animals and therefore regulate our own body temperature, all such regulation is incomplete.
When comparing 60 hours spent at 16C/60.8F to the same time spent at 22C/71.6F, rectal temperature is about 0.6C (2.52F) lower. This comes with a 6% increase in the metabolic rate.
This is when they stay in energy balance. When they overeat, they can cut the drop in core temperature in half by pushing their metabolic rate up to a 10% gain, but this still constitutes a 0.3C drop in core temperature.
In four European physicians who moved to India in 1976, prior to the spread of residential air conditioning, rectal temperatures increased from 36.8C to 37.6C after three weeks, while they were 37.8C in Indians. The fact that their temperatures rose almost as high as the Indians in just three weeks suggests that the environment was more important than genetics, and that there are no long-term adaptations to mitigate the effect of the environment.
In the winter of 1958, the arctic-adapted Norwegian Saami had lower rectal temperatures than the research team, who were from various European countries and used themselves as controls. The Saami were divided between nomadic shepherd-hunters who were accustomed to sleeping outside in the cold, protected only by tents, and villagers, who slept inside in houses. Upon arriving to the laboratory for an experiment described in the next section, the rectal temperatures were 37.3C for the researchers, 36.9C for the villagers, and 36.5C for the nomads.
The first study showed a 16C environment brought resting body temperature down to 36.4. When this is compared to the 37.8 found in Indians, it suggests at least a 1.4C spread is considered “tolerated” by the body sufficiently to avoid any adaptation.
The largest study to date on the seasonal variation in body temperature used 93,225 temporal artery temperatures taken at the point of triage at the Beth Israel Deaconess emergency department in Boston, where winter temperatures were on average 0.2C lower than summer temperatures.
In 103 soliders in Senegal, mean rectal temperatures were 0.2C higher in summer than winter.
In a single individual who took her own oral temperature for 30 years, her winter temperatures were 0.15C lower than her summer temperatures.
In ten male Japanese volunteers, rectal temperatures during one summer were 0.15-27C higher than in two different winters.
In 26 male and female subjects between the ages of 18 and 24, females had higher temperatures than males during summer months but not winter months, and there were no other statistically significant differences. The impression this gives is that temperatures were higher in summer than winter due to the jump up in female temperatures, but the statistics cannot robustly support this interpretation.
At least two studies disagree:
In 18 seasonal affective disorder patients and matched controls, rectal temperatures were slightly higher in the winter than the summer. They were taken every five minutes for 24 hours, beginning 2 hours after entry into the hospital.
In twenty men and women between the ages of 19 and 36 living in the Netherlands, rectal temperature and temperature measured with a swallowed thermometer pill indicated that core body temperature was not statistically significantly different between the end of the summer and the end of winter, but the average temperature was 0.1C higher in winter (36.9 vs 36.8). These measurements were taken after subjects stayed overnight in the lab kept at 22C and had fasted for ten hours.
My impression of these latter two studies is that having the person equilibrate to the hospital or lab’s controlled temperature is taking out the effect of the environmental temperature. If the metabolic rate goes up in winter due to incomplete compensation for the colder environmental temperature, then if you standardize the environmental temperature in which the measurements are taken, you are starting to look at the compensation, rather than the natural real-world result.
The massive dataset from the Beth Israel emergency room is vastly greater than all other data combined, and having the temperatures taken “at the point of triage” allows enough spontaneity to allow the effect of the real-world environment to assert its dominance.
In conclusion, body temperatures are about 0.2C (0.36F) colder in winter than in summer.
However, this is clearly driven by the fact that we collectively keep our homes warm in the winter.
The first study suggests that if you lower your home temperature from 22C/71.6F to 16C/60.8F to, your body temperature will drop 0.6C (1.08F), not 0.2C — providing you do not bundle up in many layers or overeat to negate the effect.
Extreme Cold Exposure Causes Long-Term Decreases in Body Temperature That Have Never Been Replicated in Cold Plunge Studies
Long-term adaptations to cold include an increased metabolic rate, and increased insulation, partly driven by increased subcutaneous fat and partly driven by unknown mechanisms. However, these never lead to complete compensation of body temperature, and adaptations to the extreme cold appear to include the body defending a lower body temperature rather than a higher one.
Whether this is due to increased cold tolerance of cells from relatively recently discovered “cold-shock proteins” or due to poorly understood redirection of heat towards certain prioritized tissues, or to changes in membrane composition to increase PUFA content and maintain optimal fluidity at lower temperatures is unclear. But, with enough stimulus from repeated heavy cold exposure, you will lower your core body temperature even further than your environment would dictate.
In the winter of 1958, as introduced earlier, the arctic-adapted Norwegian Saami were divided into villagers that slept in homes and nomads that slept in outdoor tents, and compared to European researchers who lived in then-modern housing. Upon arriving to the laboratory for a cold exposure experiment, the rectal temperatures were 37.3C for the researchers, 36.9C for the villagers, and 36.5C for the nomads. After 8 hours in a sleeping bag in a room kept at 0C, the rectal temperatures of the researchers (36.4C) and villagers (36.5C) were nearly identical, while those of the nomads dropped further to 36.0C.
This suggests that regular exposure of the nomads to the intensity of the overnight outdoor temperatures led to a specific adaptation to reach a colder core body temperature upon exposure to the same extreme cold.
The fact that the villagers did not attain any colder temperatures under these conditions than the researchers indicates that simply living in an environment cold enough to bring down your resting core body temperature does not induce long-term adaptation. The intensity of the cold found in the overnight outdoor sleeping is needed to induce long-term adaptation.
This suggests that, with sufficient purposeful training, the tolerable core body temperature can be expanded to a 1.8C (3.24F)-wide range from 36.0-37.8C (96.8-100.4F).
In 1963, the Ama diving women of Korea developed an increased metabolic rate during the winter that other Korean women did not. Their oral temperatures were not statistically significantly different at rest, though they seemed to be 0.2-0.3C higher in the colder months. However, their oral temperatures were 1.8C lower after diving shifts in January than in June, even though they were only able to dive for about 20 minutes instead of 2.5 hours.
Thus, the non-significant slightly lower resting temperature in summer could be due to spending eight times longer submerged in cold water rather than due to the higher ambient temperature. The water temperature was just slightly over 25C at the warmest time and about 10C at the coldest time. This suggests that multiple 2.5-hour exposures to 25C (77F) water are more effective than 20-minute exposures at 10C (50F)
When synthesized with the effect of the Saami nomads sleeping in outdoor tents in the arctic, it appears that considerable duration of a sufficiently intense stimulus is needed for the extra drop in body temperature.
The Ama had greater insulation, defined by less heat evolving from their body. This could not be accounted for by increased subcutaneous fat or vasoconstriction to the skin, and was speculated to be due to an unknown mechanism for evolving heat deep into their core rather than to the periphery of their body.
All of their metabolic adaptations disappeared after they started using wet suits in 1977.
In a classic study of 16 healthy men with a mean age in the mid 20s, rectal temperature was measured during 31 days of cold air exposure at 11.8C (53.2F) for eight hours per day at the end of winter and summer. At the end of winter, rectal temperature declined from 37.27 to 36.7 during the cold exposure. At the end of summer, it declined from 37.3 to 36.4 during 31 days of 13.5C. These appear to be the averages of temperatures taken at 2-hour intervals during the cold exposure, so this might reflect progressive drops in the cold-exposed temperatures rather than in the baseline temperatures. This is the only published study showing declines in cold-exposed temperatures during repeated cold exposure protocols, which suggests that the duration of eight hours per day is an important part of why some adaptation was shown.
Repeated cold exposure protocols in the literature have never been able to show a long-term reduction in resting core body temperature as found in the Saami and as seemingly found in the Ama, nor any reduction in cold-exposed temperature as shown in the study just mentioned:
In 17 male and female subjects with a mean age of 23, ten days of 2-6 hours of cold air at 15-16C (59-61F) had no effect whatsoever on resting core body temperature as measured by a swallowed thermometer pill (37.0 vs 37.0). The cold period lasted two hours on the first day, four hours the second day, and six hours thereafter. At first, the exposure period caused a 0.1C drop in core temperature but by the end it had no effect.
In 14 healthy active men with a mean age of 21, 5 weeks of daily one-hour submersions in 20C (68F) water led to an acute 0.8C drop in rectal temperature during the cold plunge but led to no change in resting temperature or response to the plunge at the end of five weeks.
Similarly, in ten young healthy men with a mean age of 22.5, 100 minutes of 10C cold air per day for ten consecutive days dropped rectal temperature by 0.4C during the cold exposure but caused no changes in resting or cold-exposed rectal temperature over the ten days.
Yet again, in seven healthy active men with a mean age of 25, a cold water adaptation protocol of 18.1C water and 21.3C air for 90 minutes over 15 days did not change resting or cold-exposed rectal temperature.
12 healthy, physically active men with a mean age of 25 were randomized to either a control group or to four 12-minute sessions per week in a 7C (44.6F) cold plunge for three weeks. Unfortunately this is the only study I can find of cold plunges with water temperatures under 10C (50F) and the only study of repeat immersion with water temperatures under 18C (64F), but it did not measure the effect on core body temperature.
Similarly, in the well-popularized study showing that cold plunging after strength training blunts hypertrophy-related adaptations in muscle, 10 minutes of a 10C cold plunge was tested twice a week over 12 weeks, making this by far the longest repeat cold plunge study, but body temperature was not measured.
Thus, while there are many potential benefits of properly dosed cold plunging for mood and cognitive function, there is zero evidence that any currently used cold plunge protocol leads to the long-term adaptations affecting core body temperature best exemplified by the Saami nomads and seemingly demonstrated by the Ama divers prior to their adoption of wet suits. Long-term adaptations to cold exposure that will lower core body temperature require intense discomfort that you are probably unwilling to experience.
It is actually much easier to lower your core body temperature with sauna.
Sauna Lowers Resting Body Temperature
In ten healthy males aged 19-22 years, 1 hour of 80C Finnish sauna at 35-38% humidity twice a day for seven days led to a linear decline in core body temperature with each successive session:

The resting rectal temperature (“before”) declined by 1.1C from 37.7 to 36.6, becoming statistically significantly different beginning on the sixth day.
The post-sauna rectal temperature, while always higher than the pre-sauna temperature, declined by 0.5C from 38.5 to 38.0, becoming statistically significantly different on the fourth day.
In 29 healthy males with mean ages in their early 20s, nine sessions of Finnish sauna at 100C and 20-30% relative humidity, each consisting of three 10-minute sauna intervals interspersed by 5-minute breaks, oral temperature appeared to decline with repeated use, although the data are slightly noisier in this study:

Focusing on “after” in set 1 of the control experimental group (EG) in the upper left, and tracing this down through each session (S1-S9), we see that temperature generally declines in each session. The ending value, however, is higher than sessions 5 and 8 but approximately the same as session 4. The session 8 value is statistically significantly lower than the session 1 value, but the session 9 value is not.
The “after” set 3 value barely moves across the sessions.
This same group did an earlier study with the same sauna protocol, except that it included a two bouts of cycling to exhaustion, one with and one without heat stress, at the beginning and end of the protocol. In this study they did not take measurements across five sessions, but only before and after the exercise bouts at the beginning and end. The ending non-heated core body temperature before exercise was slightly but non-significantly higher at the end (36.54C) than at the beginning (36.21C). However, this is probably confounded by regression to the mean, because the beginning value in the sauna group was actually statistically significantly lower than that in the control group (36.77C).
Over all, nearly all other studies performed indicate a clear decrease in core body temperature after repeated use of heat stress, but of lower magnitude than shown in the first study.
In a randomized crossover study of 13 male and female moderately fit but heat-naive participants with a mean age of 23, five days in a row of hot water immersion (HWI), sauna, or heated exercise (ExH) led to a day-by-day decrease in rectal temperature, while the regular water control (TWI) did not:

Each trial was separated by at least four weeks. The authors unfortunately do not report the absolute rectal temperatures, only the change. They also do not report whether there was a downward effect on temperature over the course of the four trials.
However, given that the magnitudes of decrease from day 1 through day 6 are similar between the trials, it is difficult to imagine that the temperatures did not recover to baseline during the four weeks between the trials.
In 27 male and female subjects between the ages of 18 and 30, those allocated to hot water immersion had their post-immersion core body temperature decrease statistically significantly by 0.2C from 38.5C to 38.3C after eight weeks of four to five sessions per week. The intervention was immersion in 40.5C water for 25-30 minutes. The authors did not report the effect on their resting core body temperature.
In 20 trained male and female runners randomized to three weeks of low-intensity outdoor running with or without 30 minutes of 101-108C sauna at 5-10% relative humidity, resting rectal temperature was not reported, but peak rectal temperature after training/sauna was 0.2C lower in the sauna group than in the control group.
Six male and female subjects with a mean age of 25 were subjected to 4 hours of sauna at 46C and 20% humidity for nine consecutive days. Their morning rectal temperature went down from 37.09 to 37.0C, but the change was not statistically significant. The afternoon rectal temperature statistically significantly declined by 0.19C from 37.17 to 36.98C.
Twelve, healthy, university-aged males were allocated (not randomly, apparently) to incremental cycling from rest to exhaustion after ten days of either sauna or control. The sauna was two hours a day of 50C at 20% relative humidity. Resting esophageal temperature declined 0.25C from 37.57C to 37.32C.
In 11 healthy males with a mean age of 25, seven consecutive days of mild heat acclimation for six hours a day at up to 37.5C led to a decrease in oral temperature of 0.12-0.14C, which was statistically significant when measured during the heating period but not in the unheated state.
In 10 recreationally active males, six days of 40-minute treadmill running at 65% of VO2max followed by up to 40 minutes of hot water immersion at 40C led to a 0.3C decline in resting rectal temperature from 37.0 to 36.7C.
13 healthy males and females with a mean age of 23 were allocated (not randomly, apparently) to a control or to seven sessions, carried out every other day for two weeks, involving 45 minutes of immersion up to the waste in 44C hot water. In the hot water immersion group, resting rectal temperature declined by 0.3C from 37.5C to 37.2C.
In six healthy males with a mean age of 23, nine days in a row of heat acclimation bringing ear temperature up to 37.5C for 100 minutes using a combination of hot water immersion and a heated blanket caused resting ear temperature to decline 0.25C from 36.76 to 36.51C.
In five healthy males with a mean age of 25, 5 days a week for just under 3 weeks of heat acclimation by keeping ear temperature at 38.5C for one hour did not appear to reduce resting ear temperature, but during any given point during heat exposure the ear temperature was about 0.5C lower after acclimation.
Synthesizing these studies, the following seem to be true:
Nearly any form of heat stress, upon repetition, causes at least a 0.2C decline in resting core body temperature.
Most studies use low temperatures in the range of 38-50C, which produces rather pathetic drops in core body temperature on the order of the 0.2C minimum.
The longest study is eight weeks and it unfortunately does not show us the time course of adaptation across the study. However, the 0.2C drop in heated core body temperature seems consistent with the magnitude of effect found in shorter studies, so it suggests that the short-term effect becomes stable and carried forward indefinitely while the stimulus is maintained.
I cannot find any observational studies comparing resting core body temperatures in long-term sauna users to controls.
By far the largest drop in core body temperature is the first study using one hour of 80C twice a day. The drop gets larger with every successive day, showing that seven days in a row is insufficient to reach the final effect.
The second study shows a much less impressive and even inconsistent drop using 30 minutes at 100C three times a week. This suggests that either 30 minutes is not long enough, that breaking the 30 minutes into 10-minute sessions with 5-minute seriously detracts from the benefit, or that three times a week is not often enough to leverage the full drop in body temperature that can be achieved.
One difference between those two studies is that the first acutely raised rectal temperature to 38.5C, while the second only raised oral temperature to 37.6C. However, the last study in the list maintained a target ear temperature of 38.5C for 100 minutes a day and generated zero drop in resting temperature. The first study kept the exposure constant, and the peak rectal temperature declined across the week from 38.5C to 38.0C. This indicates to me that the exposure should be controlled, not the body temperature reached, and that you want the sauna hot (80C) and humid (35-38%).
It is clear from the crossover study that four weeks is plenty of time to lose all of the acclimation. However, the weak and inconsistent effect in the second study suggests that much of the acclimation may be lost within 2-3 days.
The most powerful approach appears to be 1 hour of 80C twice a day at 35-38% humidity. If we define 30 minutes of 100 C three times a week at 20-30% humidity as clearly inadequate, we could say that the best range to experiment with is 30-60 minutes of 80-100C at 30-38% humidity between once every two days and twice a day.
Exercise Can But Doesn’t Always Replicate Sauna Heat, And Never Replicates the Decrease in Resting Temperature
When subjects are heated to a core temperature (measured using rectal and esophageal thermometers) of 39.5C using a 50C hot water suit, muscle temperature rises from 34.7C to 38.8C. Thus, at rest skeletal muscle temperature lags behind core temperature by over 2C, and during whole-body heating the lag becomes smaller, about 0.7C.
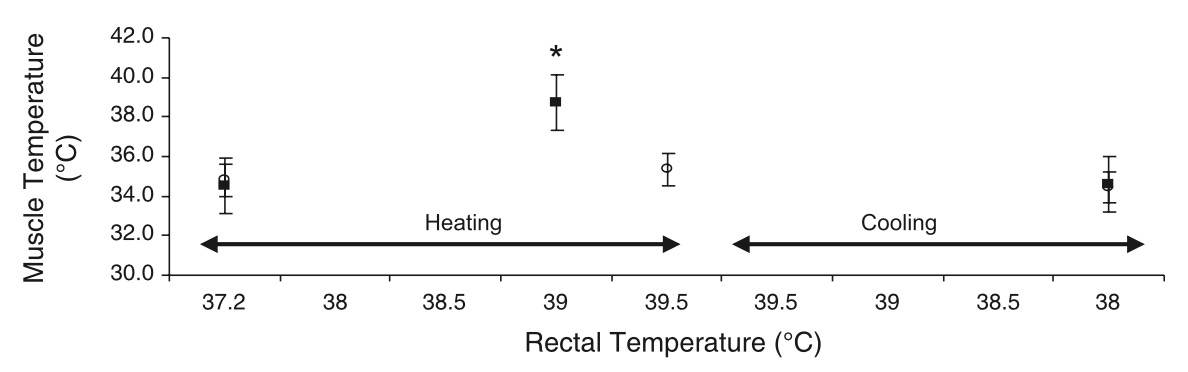
In 11 highly trained distance runners, as their running reaches close to 100% VO2max over 30 minutes, rectal temperature rises to just under 40C.
One of the subjects in that study was Edward Winrow. His rectal temperature was 40.8C after winning the 1968 Olympic Marathon Trial.
Four healthy male subjects between the ages of 21 and 32 exercised at three different intensities at three temperatures for 30-60 minutes. At 20C, after 27%, 46%, and 72% VO2max, respective rectal temperatures were 37.4, 38.0, and 38.6, while respective muscle temperatures were 38.1, 38.8, and 39.2. At 10C, rectal was 37.5, 38.0, and 38.6, while muscle was 37.9, 38.5, and 39.4. At 30C, rectal was 37.5, 38.1, and 38.8, while muscle was 38.3, 38.9, 39.9. This suggests that submaximal exercise always raises muscle temperature to 0.4-1.1C higher than core temperature, and that 72% VO2max is required to generate core temperatures similar to those achieved with sauna.
In 15 pregnant and 15 non-pregnant white Nordic female elite and recreational athletes, five five-minute treadmill intervals at about 90% of maximal heart rate interspersed by 4-minute rests raised rectal temperature to 38.4C. This suggests that high-intensity interval exercise approximates core body temperatures achieved with sauna.
In a randomized crossover trial in 12 male university Rugby players, high-intensity interval training involving 15 2-minute cycling sprints, absolute ear temperatures were not reported, but drinking an ice slurry at -2C produced a greater drop in ear temperature during rest periods than a room temperature drink, and it increased peak power. This shows that the rise in core body temperature that occurs during exercise limits performance.
12 healthy, young, physically active males with a mean age of 22 swallowed a thermometer pill and did two evening bouts each of no exercise, aerobic exercise, or resistance exercise in crossover fashion (it is unclear if it was randomized). The aerobic exercise was 30 minutes at 75% of maximum heart rate. The resistance training was three sets of ten at 75% of 10-rep max for chest press, bicep curl, seated triceps press, side lateral shoulder raise, and rear leg elevated lunge.

Core body temperature reached a sliver under 38.0 during the aerobic exercise, but only 37.5 during the resistance exercise.
As covered in Why I Don’t Care About Zone 2, when heart rate is 75% of max, percent of VO2max could be anywhere between 38% and 86%.
Thus, all exercise raises core body temperature, but neither resistance exercise at these intensities nor cardio under 90% of max heart rate and 75% of VO2 max can approximate the temperatures achieved with sauna.
In 24 adult women with chronic insomnia randomized to three sessions per week of aerobic exercise or control, rectal temperatures were nearly identical. At their highest point the exercise group was slightly higher (37.7C vs 37.6C) and at their lowest point the exercise group was slightly lower (36.3C vs 36.5C), neither of which were statistically significant.
In 24 endurance runners who had been running an average of 46 miles per week for 7-34 years, 24-hour average core body temperature measured by a swallowed thermometer pill was nearly identical (36.86C) to 24 healthy individuals consuming a western diet who were matched based on age, sex, and percent body fat (36.83C), but higher than a matched group practicing calorie restriction (36.64C).
Thus, unlike sauna, regular exercise does not seem to lower resting core body temperature.
Why Do Exercise and Sauna Differ?
It is clear that muscle temperature becomes a major driver of fatigue and one of the most important limitations of performance during exercise, so why doesn’t exercise seem to stimulate the same decrease in resting core temperature as sauna?
I suspect this is explained as follows:
During exercise, the core acts as a sink for heat generated by the muscle. While sweating is clearly used to dissipate heat out of the body, long-term adaptations could focus on moving heat out of the muscle into the rest of the body.
There are lots of other stimuli occurring during exercise and energy may be invested in many other adaptations. Further, exercise has reliable physiological triggers so the adaptations might be more specific to exercise, whereas sauna is imposed externally so the adaptations may be more generalized.
During sauna, the entire body is becoming a sink for heat generated by the environment. The only way the body can fight heat being transferred inward is to reverse this at the point of its contact with the environment.
Since the environment of a sauna is hot and humid, this imposes a very specific stress at the point of the body surface: it is harder for sweat to evaporate when the environment is humid, and it is harder to transfer heat from sweat to the environment when the environment is hot.
Thus, I believe the decrease in resting body temperature caused by sauna use is much more about the difficulty of disposing of heat at the body surface rather than about the transfer of heat within the body. The study showing the largest drop with 1 hour of 80 C at 35% humidity probably had such a large effect not because the core temperatures of the subjects were so high, but because both the heat and humidity made it hard for them to reject the heat into the environment. Thus, they rapidly adapted their ability to move heat from themselves to their environment by sweating more easily and transferring heat through sweat more easily, and when they were not in the sauna this led to a large drop in body temperature.
What About Heat Shock Proteins?
In Guatemalan sugar workers, blood levels of heat-shock proteins increase 48% by the end of the hot season. This is smaller than what can be achieved at the end of an acute bout of heated exercise, but it is actually double the rise that occurs after ten consecutive days of treadmill walking at 49C.
20 healthy males between the ages of 20 and 25 underwent ten sessions of Finnish sauna conducted over three weeks with each session being three 15-minute bouts of 90C sauna interspersed with 2 minutes of cool-down using running water three times.
The heat shock protein response was strong after the first session, but it was incredibly weak after the tenth session, and the basal levels were never meaningfully increased.

This is probably because rectal temperatures declined from 37.04C to 36.68C, indicating that the adaptation to remove heat from the body effectively trumps the need for heat-shock proteins.
This is consistent with an experiment in rats suggesting heat-shock proteins are part of the short-term rather than long-term response to heat stress. When the rats are first exposed to 41 or 43C for 2 hours, their heat shock proteins spike 4-fold to 5-fold. After two days of acclimating to a constant temperature of 34C, their basal production of heat shock proteins nearly triples. However, after 30 days of acclimation to 34C, their basal heat shock protein production goes almost to zero. When re-challenged at 41 or 43C, the proteins increase only 2-fold. Thus, for rats, during the long-term acclimation to a higher ambient temperature, other adaptations come to predominate over heat-shock proteins.
Among those, unlike humans, rats seem to be good at cooling their core body temperatures during acclimation to constant heat. Rectal temperatures increased from 37.50 to 38.04 by day one, had already fallen to 37.86 by day 2, and had fallen to 37.80 by day 30. This indicates that heat-shock proteins are mainly a function of core body temperature, and rats are better than humans at cooling their core temperatures when exposed to constant higher ambient temperature.
Thus, Guatemalan sugarcane workers have elevated HSPs after the hot season because humans are horrible at lowering our core body temperatures in response to constant heat.
However, humans are excellent are lowering our core body temperatures in response to sufficient sauna stimulus, and thus the heat-shock protein response is unneeded and practically abolished by the time the sauna stimulus has lowered our core body temperature.
Therefore, high ambient heat is bad, regular sauna use with otherwise cool ambient temperature is good, and the ability to remove heat from the body is what is important, not increasing the expression of heat-shock proteins.
Will Sauna Wreck Your Energy Metabolism?
In 1971, George Brooks and colleagues published research done on rat skeletal muscle mitochondria showing that elevated temperature from 37C to 40C increases mitochondrial respiration as a result of ATP-utilizing enzymes becoming less efficient and “leaking” energy, while temperatures between 41 and 45C appear to impair the use of oxygen to generate ATP.
In a more recent study with human natural killer cells, temperatures up to 40C had no impact at all on ATP levels. Between 41 and 45C, they started dropping off a cliff, with the depletion being stronger the higher the temperature and the longer the duration.
The ATP-depleting effects found at 41C and above would not be expected to result from sauna use. For example, the main study the sauna hard and stay cool protocol is based on had core temperatures reach 38.5C in the first session and 38.0C in the seventh session.
Human studies suggest the main impact of heat is an acute shift away from fat metabolism and toward carbohydrate metabolism.
Twelve endurance-trained males with a mean age of 22 did 40 minutes of cycling at 70% VO2 max in 20% humidity and either 20C or 40C heat in random order. Rectal temperature went from 37C to 38.6C with exercise alone, which is typical of a sauna. Exercising in heat brought rectal temperature to 39.6C, which exceeds that typically achieved by sauna. Muscle temperature reached 40.7C. These are all beneath the ATP-depleting range.
Exercise with and without heat had no effect on ATP, ADP, or their ratio; AMP was higher only after heated exercise; exercise and heat synergistically increased ammonia and the conversion of creatine phosphate to creatine; and exercise and heat synergistically and dramatically raised lactate and depleted glycogen levels.
While fat oxidation wasn’t measured, this implies an impairment of fat oxidation drove energy toward carbohydrate oxidation (glycogen depletion and lactate accumulation), amino acid oxidation (ammonia accumulation) and depletion of creatine phosphate.
This paper cites several others using higher intensities, showing that ATP was substantially depleted, so this represents effect of moderate intensity in heat, and high intensities can deplete the ATP pool (which can occur in intense exercise even without heat).
Other studies did measure carbohydrate and fat oxidation or infer their ratio.
In a randomized crossover study, 12 healthy males completed an incremental cycling test, once at 18.3C, the other at 36.3C. 21% less total fat was oxidized in the hotter environment despite no change in total energy expenditure. The total amount of carbohydrate oxidized was 26% greater but it just failed to reach statistical significance (P=0.07).
Seven male college students completed a crossover study where they cycled at 66% of maximum heart rate for sixty minutes at either 22C or -10C. Total energy expenditure was 13% greater in the cold environment, fat oxidation was 35% higher, with the carbohydrate oxidation presumably reduced to explain the difference.
A more recent study in ten healthy men with a mean age of 24 found that walking or running at 50 or 70% VO2max for sixty minutes at 0C instead of 22C caused the same shift toward fatty acid oxidation, but no difference in plasma catecholamines or free fatty acids, suggesting that the change is driven by effects on the muscle tissue itself rather than on centrally coordinated supply of substrates to the muscle via the circulation.
An experiment in rats suggests these are short-term changes. When rats are continuously exposed to 35C/95F, they deplete their glycogen acutely over 6 hours, but they recover it by 24 hours, and then they super-compensate with greater than baseline glycogen in an extended process that peaks on day 21.

As reviewed above, rats seem to adapt to constant ambient temperature rises by cooling their core temperatures, while humans do this in response to sauna but not in response to a constant ambient temperature increase.
If we use this knowledge to extrapolate from the rat study, we could infer that intermittent sauna use will acutely shift us toward carbohydrate oxidation but this will rebound after sufficient adaptation.
Two studies involving direct application of heat to muscle suggest that repeated heating leads to improvements in mitochondrial function over time.
In 20 sedentary but otherwise healthy men and women with a mean age of 21, a leg of each volunteer was randomized to serve as a control leg or to be heated with short-wave diathermy, which uses radio waves to generate heat. Heating was carried out for two hours a day for six consecutive days. Heating raised the muscle temperature by 3.9C from just under 36C to just under 40C. AMPK was acutely activated by the end of the first session. At 24 hours after the last heating session, HSP70 increased 45% and HSP90 increased 38%, and the pro-mitochondrial gene expression influencer PGC1-alpha was elevated. Complex I was increased by 37% and ATP synthase was increased by 39%, while other respiratory chain complexes were unaffected. Complex I-mediated respiration, maximal uncoupled respiration (ability to use energy to generate heat rather than ATP), and total oxidative phosphorylation were all increased 28-29%.
In 24 men and women aged 18-39, the left leg was immobilized and subjects were randomized to a diathermy or placebo treatment for two hours a day for ten consecutive days. The heat treatment raised muscle temperature from just under 36C to 40C. Immobilization had no effect on HSP70, but decreased HSP90 and PGC1-alpha. Heat increased all three of these proteins. Immobilization hurt many aspects of mitochondrial function, and heat abolished the loss of mitochondrial function caused by immobilization.
The mechanisms underlying all of this are unclear, but I would suggest the following:
If cytosolic ATP utilization becomes “leaky” the way the Brooks paper showed for mitochondrial ATP utilization, the ATP utilization in the cytosol could increase local demand for ATP recycling, putting pressure on greater glycolysis and the creatine phosphate pool to keep the cytosolic ATP pool maintained.
To the extent the recycling is biased toward the ATP demand of contracting muscle, it could come at the expense of the ATP-dependent joining of fatty acids to CoA, which is needed to enable their transport into the mitochondria for beta-oxidation.
Alternatively, or perhaps complementarily, it may be that the stability of the mitochondrial membrane is central to the carnitine shuttle. For a given temperature, this is maintained by adjusting the polyunsaturated fatty acid content (which increases fluidity) and the cholesterol content (which increases stability). Heat adds fluidity and cold adds stability. Heat therefore could hurt fatty acid oxidation by decreasing the stability of the membrane while cold could do the opposite, and this could make the cell focus on the pool of glycogen and creatine phosphate, and the collection of amino acids that can be oxidized in the cytosol, thereby completely avoiding any dependence on intracellular membrane composition. Long-term adaptation can allow cell membranes to adjust their PUFA and cholesterol content to match the fluidity needs of the repeated stress.
Difficulty meeting the demands for ATP and the massive increase in NADH that accompanies increased glycolysis drive the increases in mitochondrial complex I and ATP synthase.
Regardless of the mechanisms involved, the evidence favors that the ATP pool is not hurt by repeated sauna use, that respiratory chain function probably benefits from the hormetic stress of heat, and that fuel use is acutely biased by heat toward carbohydrate, which has a good chance of being completely counter-balanced by fat oxidation over the course of long-term adaptation.
Thus, the lower body heat that occurs with the sauna hard and stay cool protocol is driven not by decreased energy metabolism, but rather by improved removal of heat from the body.
Since muscle heat clearly limits athletic performance, one could surmise that improved removal of heat from the body is actually very helpful to energy metabolism.
In any case, the stay cool part itself should increase the metabolic rate about 6% if food intake is held constant.
Caveat: Sauna Can Probably Wreck Your Sperm Without Ice
Testicles are located outside of the body because normal body temperature is too hot for sperm.
If normal body temperature is too hot for sperm, then you would think sauna would be way too hot for sperm.
In ten healthy men with normal sperm and a mean age of 33, Finnish sauna at 80-90C and 20 to 30% humidity for 15 minutes two days a week for three months took a huge wack at sperm count and took a small bite out of progressive motility without impacting morphology of viability:

These values took six months of no sauna to go back to baseline.
A similar study using sauna for only two weeks found that sperm bounced back after only one week.
This can probably be totally avoided by keeping your testicles in an ice pack, though there is no evidence showing its efficacy. But, come on — this is why testicles are outside of the body in the first place. Just do it.
As an autistic African American who often feels colder when everyone is screaming how hot is it- I’m trying to understand this article cause it’s fascinating yet very dense.
I have my gene sequencing and such and am familiar with the genes mentioned but it’s still so much.
My husband (who is autistic and white) and I are always on opposite ends of the opinion of temperature.
So I’m going to have him help me sift through this article to understand it to apply it to us.
Thanks for sharing.
Fascinating article. It will take some time to digest but you’ve brought up so many things that pertain to me.