


The Biochemistry of Vitamin K2
The Biochemistry Underlying the Health Benefits of Vitamin K2

Vitamin K-dependent 𝛄-Carboxylation
Vitamin K2‘s best-known and most well-established role is as a cofactor for the vitamin K carboxylase. This is a role that it shares equally with vitamin K1. The difference between vitamins K1 and K2 is discussed below, and in this section I will simply refer to “vitamin K.”
The vitamin K carboxylase is an enzyme that adds carbon dioxide to the side chains of specific glutamate residues within specific vitamin K-dependent proteins. Once added to a glutamate residue, the carbon dioxide becomes a carboxyl group, so the process is known as carboxylation. Carboxyl groups carry negative charges, so carboxylation helps vitamin K-dependent proteins bind to calcium, which carries a positive charge. In other words, the most well-established role of vitamin K is to add carbon dioxide to proteins and thereby give them the ability to bind calcium.
Since the carboxyl group is added to the third carbon of the glutamate sidechain, known as the gamma (𝛄) carbon, the process is known as 𝛄-carboxylation. Once modified in this way, glutamate becomes 𝛄-carboxyglutamate and is abbreviated “Gla.” Thus, vitamin K-dependent proteins often have the term “Gla” in their names.
The figure below shows vitamin K-dependent 𝛄-carboxylation in more detail.
A. The general structure of an amino acid, featuring a carboxyl group (COOH) on the right.
B. At the pH range that prevails within the human body, carboxyl groups often ionize, giving them a negative charge.
C. Glutamate. Moving from the central carbon through each carbon of the side chain, we label the carbons alpha (α), beta (β), and gamma (ɣ). Since the side chain carboxyl group is attached to the ɣ carbon, it is known as a ɣ-carboxyl group.
D. ɣ-carboxylation. The vitamin K carboxylase uses vitamin K as a cofactor to add carbon dioxide to the ɣ carbon of the glutamate residue side chain. This converts glutamate, abbreviated Glu, to ɣ-carboxylglutamate, abbreviated Gla. The second ɣ-carboxyl group gives the side chain a second negative charge, which improves its ability to bind to calcium, which carries a positive charge. Although ionic calcium is shown in the figure, some vitamin K-dependent proteins bind to calcium salts rather than calcium ions.
There are a wide variety of vitamin K-dependent proteins made in different tissues that fulfill different functions but that all undergo the same process of 𝛄-carboxylation. In each case, calcium-binding is essential for the protein’s activities. However, the role that calcium-binding plays is different for different proteins.
The Role of Vitamin K in Blood Clotting
We have known about the importance of vitamin K for blood clotting since the 1930s (Suttie, 2014). Blood clotting is regulated by a variety of proteins known as clotting factors that are all made in the liver and sent out into the blood, where they circulate in inactive form until blood vessel damage makes clotting necessary. In the clotting factors, vitamin K-dependent 𝛄-carboxylation allows calcium to serve as a structural “glue” that binds the protein into an active shape.
Initially, we only knew that vitamin K was necessary to the function of prothrombin, the precursor to thrombin, which activates fibrinogen to fibrin to form blood clots. We now know that vitamin K is also needed for properly functioning factors VII, IX, and X, which are pro-coagulant proteins involved in the conversion of prothrombin to thrombin. Vitamin K is just as necessary for the function of proteins S and C, which act as anticoagulants by inactivating other clotting factors that are not dependent on vitamin K, factors V and VIII. There is a seventh vitamin K-dependent plasma protein, protein Z, that may have additional anticoagulant functions. Thus, vitamin K is required for the proper function of both procoagulants and anticoagulants within the clotting cascade and serves as a raw material necessary for the proper regulation of blood clotting rather than serving as a coagulant or an anticoagulant itself.
The Role of Osteocalcin in Metabolic and Hormonal Health
Vitamin K is necessary for the carboxylation of osteocalcin, a protein produced in bone and also sometimes referred to as bone Gla protein. In this case, vitamin K-dependent 𝛄-carboxylation allows osteocalcin to bind to the calcified extracellular matrix of bone tissue (Koshihara, 1997).
Scientists first discovered osteocalcin in the 1970s. Since it was made in bone, most scientists in the field assumed that it played an important role in mineralizing bone or in regulating the turnover of bone mineral or the structural organization of bone. In the 1990s, however, scientists produced the osteocalcin-knockout mouse, which is genetically modified to lack the gene that codes for osteocalcin. Osteocalcin knockout mice have no obvious defects in any measures of bone health. Their bones are adequately mineralized, and although their structural architecture is slightly different than that of normal mice, one study found their bones were stronger than those of normal mice (Ducy, 1996) and the worst that could be said about their bone architecture was that it seemed “less mature” than that of normal mice (Boskey, 1998).
For three decades, the role of osteocalcin was elusive and the statements made about its function were vague and unconvincing.
In 2007, things began to change (Lee, 2007). The scientists who had developed the osteocalcin knockout mouse began more intensively investigating their phenotype and publishing papers about their metabolic and hormonal health. And here, unlike in bone, the effects of osteocalcin are dramatic.
Osteocalcin knockout mice are fat, deficient in insulin (like type 1 diabetics), insensitive to insulin (like type 2 diabetics), and they have low metabolic rates and high blood sugar. The males are also infertile and have low testosterone (Oury, 2011).
Surprisingly, all of this is reversed with undercarboxylated osteocalcin rather than fully carboxylated osteocalcin. Undercarboxylated osteocalcin is produced by bone when vitamin K status is inadequate, and its circulation in serum had been interpreted as a sign of vitamin K inadequacy right up through the publication of these papers. In fact, some vitamin K researchers argue that it should still be used in this way, adding controversy to the implications of the osteocalcin knockout mouse studies (Booth, 2013).
I believe the best way to reconcile these conflicting ideas is as follows: vitamin K-dependent 𝛄-carboxylation of osteocalcin takes place in osteoblasts and allows the carboxylated osteocalcin (cOCN) to leave the osteoblasts and accumulate in bone matrix, which is its proper site of storage. During bone resorption, osteoclasts produce acid that decarboxylates osteocalcin and releases it into the serum in its undercarboxylated form (ucOCN) (Ferron, 2007; Oury, 2013). From there, it acts on multiple tissues to improve insulin secretion, insulin sensitivity, blood glucose, the metabolic rate, body composition, and, in males, testosterone production and fertility. This is illustrated in the figure below.
Most recently, the release of undercarboxylated osteocalcin from bone was shown to increase during exercise and play a role in allowing skeletal muscle to increase its utilization of energy (Mera, 2016). Exercising skeletal muscle secretes interleukin-6 (IL-6), which increases the release of undercarboxylated osteocalcin (ucOCN) from bone. ucOCN stimulates muscle to release more IL-6, and they amplify one another in a positive feedback loop. IL-6 acts on liver to release glucose and adipose tissue to release free fatty acids. IL-6 and ucOCN act on skeletal muscle to increase the uptake of glucose and fatty acids and increase their utilization for energy. This is illustrated in the figure below.
Matrix Gla Protein (MGP) Regulation of Calcium Distribution
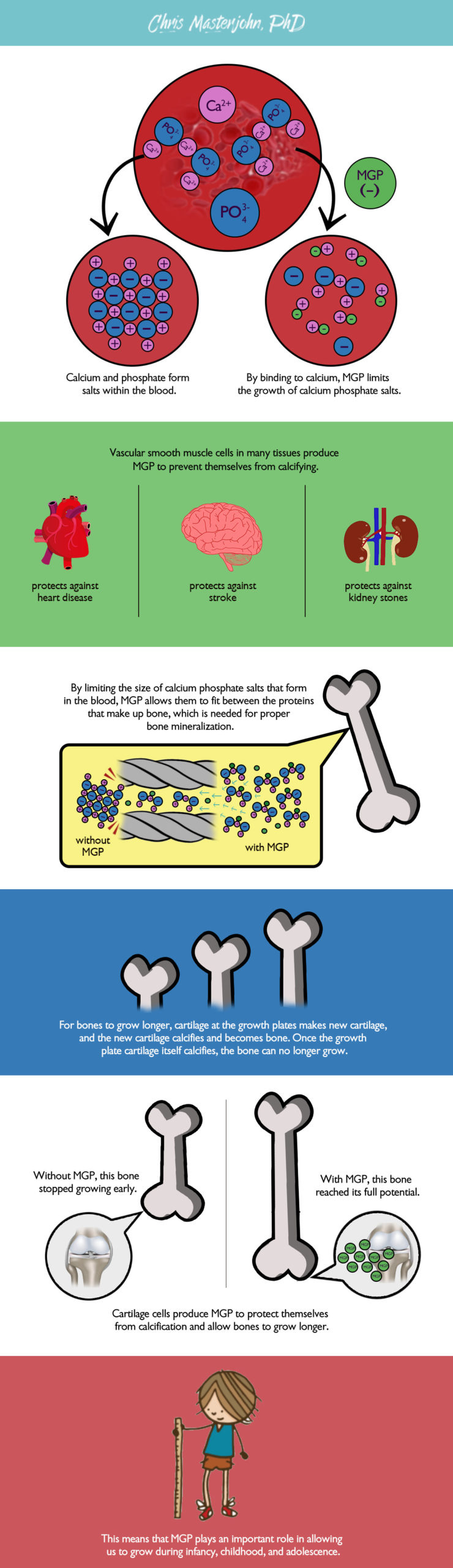
Vitamin K is necessary for the carboxylation of matrix Gla protein (MGP), which is made primarily in vascular smooth muscle cells and chondrocytes (cartilage cells) (Luo, 1997). Wherever there is a blood supply, there is MGP, so MGP is made throughout the body. In this case, vitamin K-dependent 𝛄-carboxylation allows MGP to bind calcium so that it can prevent calcium from going into the wrong places, like into the kidneys and blood vessels, and help it go into the right places, like the extracellular matrix of bones and teeth.
MGP appears to act primarily by limiting the formation of calcium salts. This helps prevent pathological calcification of soft tissues (tissues other than the bones and teeth). For example, MGP protects against kidney stones and against the calcification of blood vessels that occurs in heart disease. Bone has a complex protein infrastructure that becomes mineralized through the entry of very small calcium phosphate salts from blood. By limiting the size of these salts (Price, 2009), MGP helps them penetrate bone matrix and support its mineralization. MGP also supports growth during infancy, childhood, and adolescence by preventing premature calcification of the cartilage that helps bones to become larger.
These roles of MGP are illustrated in the figure below.
MK-4 and Gene Expression
As described below, different forms of vitamin K reach different tissues to different degrees, so some forms better support some of the health outcomes discussed above than others. However, all the roles described above can be fulfilled by any form of vitamin K able to reach the relevant tissues. By contrast, MK-4 is a subform of vitamin K2 that has a unique role in regulating gene expression (Ichikawa, 2007; Ito, 2011). The mechanisms involved are unclear: some studies show that it binds to the steroid X receptor (SXR), while others show that it regulates gene expression through SXR-independent mechanisms. One of those mechanisms is to stimulate the phosphorylation of protein kinase A (PKA), but how it does this is also unclear: some studies show that it increases cyclic AMP (cAMP), a traditional PKA activator, while other studies show it activates PKA independently from cAMP. In other words, we know that MK-4 regulates gene expression, but we have a lot to learn about how it does this.
Through its regulation of gene expression, MK-4 favors bone growth, protects against cancer, and increases the production of sex hormones.
Other Functions of Vitamin K
There are a variety of other vitamin K-dependent proteins whose functions are less clearly understood (Suttie, 2014). These include the following: Gla-rich protein, which accumulates in soft tissues during pathological calcification; periostin, which may be necessary for growth; Gas6, which promotes cell survival, and, along with protein S, helps clear away the debris of dead cells (for example, in atherosclerosis, where accumulating debris of dying cells causes a dangerous inflammatory state); and a family of four transmembrane Gla-rich proteins may act as cell surface receptors. Vitamin K also supports the production of important sulfur-based lipids known as sulfatides in the brain, and accumulates in the mitochondrion where it may play a direct role in the electron transport chain, as it has been shown to do in fruit flies (Vos, 2012).